Биологическая сущность гетерозиса. Понятие, типы, значение гетерозиса Явление гетерозиса и его цитологические основы
Под гетерозисом понимают превосходство потомства первого поколения над родительскими формами (лучшей родительской формой) по жизнеспособности, конституциональной крепости, выносливости и продуктивности, возникающее при скрещивании разных рас, пород животных и их зональных типов (Е.К. Меркурьева и др., 1991 ).
Некоторые ученые считают эффектом гетерозиса и те случаи, когда потомство, полученное в результате скрещивания, имеет показатели хозяйственно полезных признаков, превышающие средние между родительскими формами, не превосходя лучшие из них.
Селекция на получение гетерозиса непосредственно связана с теорией и практикой племенного отбора и подбора и служит одним из путей повышения продуктивности животных. Родственное спаривание сопровождается инбредной депрессией, повышением гомозиготности инбредного потомства и увеличением генетического сходства потомка с предком. Противоположными биологическими и генетическими свойствами обладает гетерозис.
Основными показателями гетерозиса являются повышение эмбриональной и постэмбриональной жизнеспособности; снижение затрат корма на единицу продукции; повышение скороспелости, плодовитости, продуктивности; проявление более широких возможностей приспособления к смене условий и новым элементам технологии. Широкий диапазон эффекта гетерозиса, проявляющийся в многообразии реагирующих признаков, является отражением физиологических и биохимических процессов, обусловленных особенностями генетического аппарата гетерозисных животных. Одной из особенностей его является наибольшая степень выраженности лишь в 1 поколении гибридов или помесей. Затем гетерозис незаметно затухает и исчезает в следующих поколениях при скрещивании гибридов друг с другом, если не принимается специальных мер для сохранения эффекта гетерозиса.
Явление гетерозиса, или "гибридной силы", было замечено в практике Животноводства в давние времена, в частности при получении мулов скрещиванием осла с кобылой. Мощнее исходных форм получаются также гибриды от двугорбого и одногорбого верблюдов (батриан+дромедар= нары и майя), от яка и коровы (хайнаки).Ч. Дарвин впервые дал научное объяснение "гибридной силы", которая возникает у потомства при скрещивании неродственных организмов. Он объяснял этот эффект биологическим несходством мужских и женских гамет, которое вызывается влиянием различий окружающей среды, в которой обитают родители.
Термин "гетерозис" был введен Г. Шеллом (1914), который объяснял наличие "гибридной силы" состоянием гетерозиготности в генотипе организма, формирующейся в результате скрещивания.
Попыток вскрыть механизм самого процесса сочетаемости наследственных факторов, обусловливающего эффект гетерозиса, было много. Шелл и Ист предложили гипотезу сверхдоминирования . Гипотеза гетерозиса, сформулированная Шеллом, Истом и Хейсом, объясняет явление гетерозиса наличием гетерозиготности различных локусов и проявляющимся при этом сверхдоминированием, то есть когда действие гетерозиготы Аа на проявление фенотипа оказывается сильнее, чем гомозиготного доминантного генотипа АА (то есть эффект действия Аа больше действия АА). Т.о. суть ее заключается в том, что высокая гетерозиготность лучше, чем гомозиготность, обеспечивает разнообразие и усиление физиологических функций организма. Значение гетерозиготности было подтверждено работами Н.П. Дубинина, М. Лернера и других ученых.
Другое объяснение гетерозиса, сформулированное Кийблом и Пеллью (1910), основано на том, что при скрещивании организмов, несущих в генотипе разные гомозиготные гены, например ААbb и ааВВ, у помесного потомства рецессивные аллели переходят в гетерозиготную форму генотипа АаВb, при которой устраняется вредное действие рецессивных генов. Влияние доминантных генов на проявление гетерозиса может быть объяснено простым суммарным действием большого количества доминантных генов, то есть имеет место аддитивный эффект.
К. Давенпорт (1908) и Д. Джонс (1917) предложили объяснять гетерозис исходя из гипотезы взаимодействия неаллельных доминантных генов обоих родителей, что дает суммарный эффект, вызывающий гетерозис. Кисловский Д.А. для объяснения гетерозиса выдвинул гипотезу облигатной гетерозиготности , согласно которой в организме имеются гены с двойным действием - полезным и вредным. В одном направлении это действие полезно, в другом нейтрально или даже вредно для организма. В процессе эволюции выживают те организмы, у которых положительное действие генов выявилось в гетерозиготном состоянии, а вредное оказалось в рецессивном. Гетерозис прежде всего полезен самому организму животного, происходящему от скрещивания. Отсюда следует, что высокая степень гетерозиготности - причина гетерозиса .
Для объяснения явления гетерозиса в последние годы была выдвинута зоотехническая концепция гетерозиса (А.И. Овсянников, И. Н, Никитченко и другие ) суть которой заключается в следующем:
- 1. Концепция контрастных скрещиваний. Согласно ей, эффективность скрещивания связывается в основном с контрастными и противоположными по направлению и типу телосложения родительскими парами. Принцип подбора пар для получения гетерозиса основан на проверенном эффекте гетерогенного спаривания маток с отдельными признаками продуктивности в сочетании с ценными признаками отца. Сочетание редких крайностей не допускается (идет комплексный подбор). Повышение жизнеспособности, гибридной силы должно достигаться подбором пород и особей, различных по экстерьеру, обмену веществ, интерьерным показателям.
- 2. Принцип дополняющего действия. Ведущая роль в формировании гетерозиса принадлежит сочетающейся различиями наследственности исходных пород. При этом определено, что гетерозис представляет собой сложное биологическое явление, в котором решающее значение имеют 4 группы факторов:
- а) прямое действие генов (уровень и число продуктивных качеств исходных пород);
- б) материнский (реципрокный) эффект;
- в) дополняющее действие генетических факторов (аддитивное), действие доминантных генов, накопление которых в потомстве при скрещивании усиливают развитие признака, вызывая гетерозис;
- г) условия жизни приплода I поколения.
Практикой апробированы различные методы получения гетерозисных животных. К ним относятся: межвидовые скрещивания, межпородные скрещивания, внутрипородные скрещивания при гетерогенном подборе, межлинейные кроссы, кроссы специально создаваемых инбредных линий, спаривание животных, выращенных в различных условиях. Каждый из этих методов имеет свои особенности и может быть использован для получения гетерозиса не по всем, а лишь по определенным признакам. Какие бы методы не использовали для получения гетерозиса, большое значение имеют индивидуальные особенности производителя. Чем ценнее его происхождение и выше способность передавать свои качества потомству, тем при прочих равных условиях будет выше степень проявления гетерозиса.
Огромная роль гетерозиса в повышении продуктивности и улучшении других хозяйственно-полезных признаков животных побуждала многих ученых изыскивать пути длительного закрепления его или хотя бы сохранения в течение нескольких поколений. Д.А. Кисловский одним из первых теоретически обосновал возможность использования гетерозиса в последующих поколениях при межпородном переменном скрещивании. Он утверждал, что при таком скрещивании как бы комбинируются особенности и положительные стороны поглотительного и промышленного скрещиваний.
Существуют и другие биохимические объяснения гетерозиса . Считается, что главной причиной гибридной мощности служит формирование на хромосомах чувствительных копий структурных генов, которые образуют избыток информации в клетках и определяют высокую сочетаемость процессов метаболизма (Северин, 1967 ). Объяснения гетерозисному эффекту можно найти в суждениях, что у помесей присутствуют полиморфные типы белков (изоферменты), которые различаются некоторыми свойствами. У родительских форм отсутствует полиморфизм ферментов, а при их скрещивании у помесей формируется полиморфизм и число полиморфных локусов у них поэтому больше, чем у родителей. Это, по мнению некоторых ученых (Финчем, 1968; Кирпичников, 1974 ), объясняет эффект сверхдоминирования.
Ф.М. Мухаметгалиев (1975) считает, что взаимное стимулирование геномов при оплодотворении равносильно аддитивному эффекту объединенных генетических систем и является основой появления гетерозиса, но не является причиной возникновения новых качеств в генетическом материале, поэтому гетерозис проявляется в количествнных изменениях признаков и имеет полигенный тип наследования.
Новый подход в объяснении гетерозисного эффекта предлагает В.Г. Шахбазов (1968). Он считает, что гетерозис имеет биофизическую основу, так как при оплодотворении происходит обмен электрическими зарядами гомологичных хромосом, что повышает активность хромосом в гибридных зиготах. Это приводит к накоплению кислых белков и РНК, повышает ядрышко-ядерное соотношение и увеличивает скорость митотического деления.
Приведенные объяснения причин гетерозисного эффекта указывают на отсутствие единства в научном объяснении явления гетерозиса, и поэтому проблема остается для дальнейшего изучения и рассмотрения. Несмотря на это, в практике животноводства осуществляют приемы селекции животных на закрепление и усиление эффекта гетерозиса. Существует несколько приемов для вычисления величины эффекта гетерозиса. Выделяют так называемый истинный тип гетерозиса , который определяется по величине превосходства признака у помесных животных над обоими родительскими формами. Другой тип гетерозиса - гипотетический , когда признаки помесного потомства превосходят среднеарифметический уровень признака обоих родителей.
Профессор Н.А. Юрасов определяет линию как микропороду. С этой точки зрения удачный кросс линий можно рассматривать как результат проявления гетерозиса. Подобно тому, как гетерозис при межпородных скрещиваниях является результатом генетического разнообразия родителей, так и гетерозис при кроссах линий можно объяснить тем же явлением. Чем тщательнее селекционированы и консолидированы кроссируемые линии, тем более высокий эффект можно ожидать.
Испытание родительских пар и целых линий на сочетаемость для получения гетерозиса лежит в основе одного из эффективных приемов, который получил название непрерывной реципрокной селекции . Современные представления о причинах появления гетерозиса основаны на том, что гетерозис является результатом взаимодействия многих генов. Их множественное действие и приводит к гетерозисному эффекту. Такое объяснение получило название балансового гетерозиса (Добжанский, 1952 ).
Выявлен экологический тип гетерозиса (Меркурьева, 1980 ), который вызывается процессом акклиматизации и проявляется у животных первой экологической генерации. Этот тип гетерозиса проявился в повышенной молочности потомства, родившегося в Рязанской области от айрширских коров, завезенных из Финляндии. В последующих поколениях удои снижались до уровня, соответствующего генетическому потенциалу завезенной группы коров.
С целью специального использования гетерозиса в зоотехнии разработаны методы промышленного и переменного скрещивания. В таких случаях скрещивают чистопородных животных двух пород, а их потомство, превосходящее представителей исходных пород, целиком или в большей части используют для откорма и забоя на мясо. Больше всего примеров проявления гетерозиса у животных разных видов отмечено при межпородных скрещиваниях. В мясном скотоводстве при скрещивании разных пород помеси I поколения превосходят исходные породы по живой массе. У молочного скота эффект гетерозиса наблюдается чаще по общему количеству молочного жира за лактацию, особенно при скрещивании коров различных пород с быками джерсейской породы.
При скрещивании некоторых сортов какой-либо культуры гибриды F1 часто отличаются от родительских форм более мощным ростом, лучшей жизнеспособностью, более высокой продуктивностью, устойчивостью к болезням и резким колебаниям погоды. Такое свойство гибридов первого поколения называют гетерозисом.
Гетерозис - это свойство гибридов первого поколения превосходить родителей или лучшую из родительских форм по определенным биологическим и хозяйственно ценным признакам и свойствам, по степени их выраженности.
Проявление гетерозиса зависит от вида генофонда популяции, физиологического состояния популяции на определенном этапе развития, от условий климата, сезона, солнечной инсоляции, питания, густоты посева или посадки, напряженности инфекционного фона или насекомых вредителей или других факторов и обстоятельств.
Истинный (положительный) гетерозис проявляется в изменении выраженности признака у гибрида в сторону увеличения по сравнению с родительскими формами (например, повышенное содержание витаминов, белков, биологически активных веществ, улучшение лежкости продукции и др.). Однако при скрещивании могут возникнуть депрессивные явления снижения потомством показателя по какому либо признаку лучшего родителя. Такое изменение выраженности признака в сторону ослабления происходит при отрицательном гетерозисе.
Гетерозис может охватывать разные признаки и свойства организма. А. Густафсон предложил различать три основные типа гетерозиса: соматический, репродуктивный и адаптивный. Гетерозис соматический - более мощное развитие вегетативных органов гибридов; репродуктивный - более мощное развитие репродуктивных органов, повышенная фертильность, высокий урожай семян, плодов; адаптивный гетерозис - повышение приспособленности гибридов к меняющимся условиям среды и их конкурентоспособность в борьбе за существование.
Гетерозисные гибриды первого поколения F1 проявляют повышенную жизнеспособность, мощность развития, особо сильную способность к росту, большую скороспелость, урожайность и выравненность, повышенную устойчивость к болезням, высокую товарность, улучшение качества плодов (повышенное содержание сухого вещества).
Гетерозис может проявляться в изменении физиологических признаков - усилении холодостойкости, засухоустойчивости, улучшении лежкости, общей устойчивости к патогенам.
Гетерозис проявляется в более быстром росте растений в начальных и последующих фазах, в ускоренном образовании новых листьев, в увеличении размера каждого листа и общей площади листовой поверхности, в мощности корневой системы и др. гетерозис проявляется также в таких признаках, как высота закладки соцветий, количество соцветий на главном стебле и боковых побегах, количество цветков в соцветии и т.д.
Гетерозис наблюдается при скрещиваниях между сортами, а также между отдаленными в генетическом и экологическом отношении видами и формами. Наиболее же сильно он проявляется и поддается управлению при скрещивании самоопыленных линий. Инцухт дает возможность разложить сорт-популяцию на составляющие его биотипы (линии).
Проявление гетерозиса наблюдается в течение всего жизненного цикла растения с момента до окончания формирования семени.
Тот или иной тип проявления гетерозиса и степень изменчивости выражения признака при этом в основном зависят от подбора скрещиваемых сортов. Обычно гетерозис бывает тем сильнее выражен, чем больше различаются скрещиваемые формы по морфологическим, биологическим, физиологическим и другим признакам. Особенно сильно проявляется гетерозис, если скрещиваемые сорта относятся к разным группам по скороспелости, способу выращивания, если районированы они в разных климатических зонах или культивируются в разное время года, при скрещивании инбредных линий перекрестноопыляющихся растений, полученных в результате самоопыления в течение нескольких поколений .
Одной из причин эффекта гетерозиса является устранение в гетерозиготном организме вредного действия рецессивных генов. Другой механизм гетерозиса состоит в объединении в гибриде благоприятных доминантных генов, присутствующих в организме родителей и объединяющихся у гибрида. Гетерозис может возникнуть и в силу того, что некоторые гены более благоприятно проявляются, находясь в организме в гетерозиготном состоянии.
ЗНАЧЕНИЕ СЕЛЕКЦИИ НА ГЕТЕРОЗИС
Селекцией на гетерозис называют создание гибридов первого поколения, отличающихся высоким гетерозисом по урожайности, качеству продукции и другим хозяйственно важным признакам. В отличие от комбинационной селекции, при которой скрещивания проводят в начале селекционного процесса, чтобы создать генетическую изменчивость для отбора, при селекции гетерозисных гибридов скрещивание служит для массового получения семян и их дальнейшего практического использования в производстве и представляет последний этап селекционного процесса. Селекция гетерозисных гибридов имеет большое значение для сельскохозяйственного производства. Эти гибриды часто по урожайности превышают обычные свободноопыляющиеся сорта на 30% и выше. В некоторых случаях гетерозисный эффект достигает 50%.
В селекции животных и растений особое место занимает явление гибридной мощности, или гетерозиса , которое заключается в следующем.
При скрещивании разных рас, пород животных и сортов растений, а также инбредных линий гибриды F 1 по ряду признаков и свойств часто превосходят исходные родительские организмы. Скрещивание гибридов между собой ведет к затуханию этого эффекта в следующих поколениях. В настоящее время гетерозис установлен для всех изученных видов.
Хотя эффект гетерозиса известен с древнейших времен, его природа до сих пор остается мало изученной. Первое толкование биологического значения гетерозиса в эволюции животных и растений и попытка объяснить механизм этого явления принадлежали Ч. Дарвину. Он систематизировал большое количество фактов и сам ставил опыты по изучению гетерозиса. По мнению Ч. Дарвина, гетерозис служит одной из причин биологической полезности скрещивания в эволюции видов. Перекрестное оплодотворение поддерживается естественным отбором именно потому, что оно служит механизмом сохранения наибольшей гетерозисности,
Глубокий научный анализ явления гетерозиса стал возможен только с начала XX в. после открытия основных генетических закономерностей. При рассмотрении гетерозиса мы будем касаться возникновения его при скрещивании преимущественно инбредированных линий одного вида, поскольку в этом случае легче выяснить его генетический механизм.
С начала нашего столетия на кукурузе стали проводить систематическое исследование скрещиваний между инбредными линиями. При этом Г. Шеллом было показано, что скрещивание некоторых линий дает гибридные растения, более урожайные по зерну и вегетативной массе, чем исходные линии и сорта. Приведены опытные данные, показывающие низкую урожайность инбредных линий, значительное повышение урожайности в F 1 и снижение в F 2 при самоопылении растений F 1 .
За последние 30 лет в ряде стран, в том числе и в нашей, посев гибридными семенами стал основным приемом производства кукурузы как на зерно, так и для силоса. Для получения гибридных семян сначала создают инбредные линии из лучших сортов, отвечающих требованиям данного климатического района (инбредная линия создается в течение 5-6 лет путем самоопыления). При отборе линий оценивают их качества и свойства, которые необходимо получить у будущего гибридного организма. Инбридинг в линиях не может быть эффективным, если он не сопровождается отбором.
Создав большое число инбредных линий, приступают к скрещиванию между ними. Межлинейные гибриды первого поколения оценивают по эффекту гетерозиса ; исходя из этого показателя отбирают линии с лучшей комбинационной ценностью и затем размножают их в больших масштабах для производства гибридных семян. Работа по созданию инбредных линий и оценке их комбинационной ценности постоянно ведется в селекционных учреждениях. Чем больше создано ценных линий, тем вернее й скорее можно отыскать лучшие гибридные комбинации с необходимым сочетанием свойств.
При получении гибридных семян для производственных целей исходные линии, дающие при скрещивании наибольший эффект гетерозиса, высевают рядами, чередуя материнские и отцовские формы. Чтобы обеспечить опыление между ними, с материнских растений удаляют мужские соцветия (метелки). Теперь разработана новая схема производства гибридных семян с использованием цитоплазматической мужской стерильности, что позволило значительно сократить труд на удаление метелок у растений материнской линии. Таким путем получают простые межлинейные гибриды кукурузы. Этот метод в принципе является общим для семеноводства гибридов различных перекрестноопыляющихся растений.
В настоящее время в практике сельского хозяйства простые межлинейные гибриды кукурузы не используются, так как затраты на получение таких семян не окупаются. Теперь широко внедряется в практику посев семян двойных межлинейных гибридов. Последние получают путем скрещивания двух простых гибридов, проявляющих гетерозис.
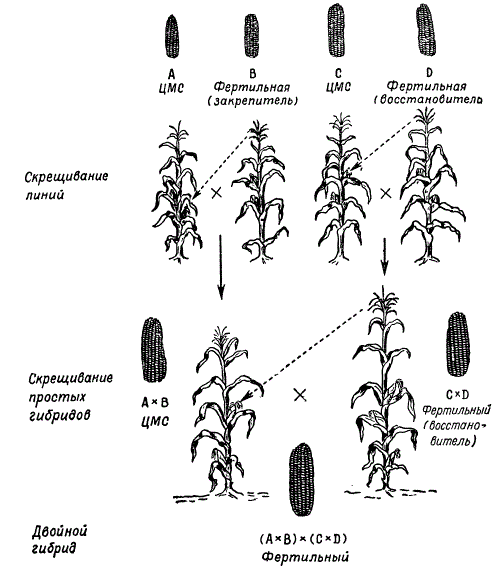
По данным М. И. Хаджинова и Г. С. Галеева, результаты оценки продуктивности разных гибридов - межсортовых, сортолинейных и двойных межлинейных - показывают, что наиболее продуктивны двойные межлинейные гибриды.

Подбор простых гибридов для получения наиболее продуктивных двойных гибридов является важным этапом их селекции. Лучшие результаты дает скрещивание линий, происходящих из различных сортов. Так, например, если один простой гибрид получен от скрещивания инбредных линий двух сортов А X В, а другой - от скрещивания линий других сортов С х D, то двойной гибрид (А X В) х (С X D) дает гетерозис чаще, чем если бы двойной гибрид был получен от скрещивания простых гибридов, происходящих от линий одного сорта: (А х А 1 х (А 2 х А 3) или (В х B 1) х (В 2 х В 3).
Для успешного развития гибридного семеноводства необходимо исследовать, во-первых, как долго следует вести инбридинг, чтобы получить гомозиготные линии, и, во-вторых, разработать методы более быстрой оценки их комбинационной ценности.
Все, что было сказано в отношении кукурузы, по-видимому, является общим для получения гибридов у других высших перекрестноопыляющихся растений, а также у животных. В настоящее время в птицеводстве и в свиноводстве ряда стран широко используется скрещивание инбредных линий, происходящих из одной или разных пород. Следует особо подчеркнуть, что широкое использование гибридов в животноводстве возможно только при высоком уровне племенной работы и наличии ценных пород, то совершенно ясно, что в большинстве случаев инбредные линии в среднем будут иметь всегда более низкие показатели, чем сорта. О наличии гетерозиса следует говорить лишь в том случае, когда межлинейный гибрид превосходит не только родителей (линии), но и сорта, или породы, от которых произошли эти линии.
Как мы знаем, гены детерминируют свойства организма на всех стадиях онтогенеза начиная с момента оплодотворения. Гены ядра ооцита способны детерминировать свойства цитоплазмы яйцеклетки еще до оплодотворения. Характер реализации генотипа изменяется в зависимости от свойств цитоплазмы зиготы. Таким образом, проявление гетерозиса у гибрида зависит также и от свойств цитоплазмы. Роль цитоплазмы в определении гетерозиса доказывается следующим образом. При реципрокном скрещивании двух линий А X В и В х А гетерозис по одним и тем же свойствам нередко проявляется у гибридов только одного из скрещиваний и не проявляется у гибридов другого.
Проявление гетерозиса обусловлено индивидуальным развитием гибрида. В онтогенезе он реализуется неравномерно. На одних стадиях онтогенеза проявляется гетерозис по одним признакам, а на других - по другим или только по некоторым. Так, в раннем возрасте у одного и того же гибрида может наблюдаться гетерозис в отношении скорости роста отдельных частей организма и повышенной устойчивости к заболеваниям, но его может не быть, например, в отношении устойчивости к неблагоприятной температуре. Гетерозис по этому свойству может проявляться позднее.
Сильное влияние на проявление гетерозиса оказывают также факторы среды, в которых развивается гибридный организм. Есть основание считать, что гетерозис по жизнеспособности и устойчивости к заболеваниям лучше выявляется при неблагоприятных для развития условиях среды. Еще Ч. Дарвином было высказано предположение, что гетерозис у гибридов обусловлен более широкой нормой приспособительных реакций. Исследования последних лет показали, что гибриды F 1 в сравнении с инбредными линиями характеризуются большим разнообразием процессов обмена, участием большего числа различных метаболитов, ростовых веществ и энзимов. Гетерозис проявляется не только в целом организме, но и на клеточном уровне.
Выяснение генетических механизмов гетерозиса остается нерешенной проблемой. В настоящее время имеются три гипотезы, пытающиеся объяснить возникновение гетерозиса:
- гетерозиготное состояние по многим генам,
- взаимодействие доминантных благоприятных генов,
- сверхдоминирование - гетерозигота превосходит гомозиготы.
Как мы уже говорили, при скрещивании гомозиготных инбридированных линий гибриды первого поколения по многим генам приобретают гетерозиготное состояние. При этом действие вредных рецессивных мутантных аллелей подавляется доминантными аллелями обоих родителей. Схематически это можно представить так: одна инбредная линия в гомозиготном состоянии имеет рецессивную аллель одного гена ааВВ, а вторая - другого гена ААbb. Каждая из этих рецессивных аллелей генов в гомозиготном состоянии определяет какую-нибудь недостаточность, которая снижает жизнеспособность инбредной линии. При скрещивании линий ааВВ X ААbb у гибрида объединяются доминантные аллели обоих генов (АаВb). Гибриды F 1 проявят при этом по указанным генам не только гетерозис, но и единообразие. В F 2 число особей с двумя доминантными генами в гетерозиготном состоянии будет лишь 4/16, поэтому гетерозисными оказываются не все особи. В дальнейших поколениях число гетерозигот сокращается, а число гомозигот увеличивается. В силу этих причин гетерозис в последующих поколениях затухает. Такова схема одной из генетических гипотез гетерозиса.
Мы привели пример с двумя генами, но физиологические свойства определяются огромным числом генов. Кроме того, неверно считать, что доминантные аллели всегда положительно, а рецессивные отрицательно влияют на жизнеспособность организма и его продуктивность. Доминантные аллели дикого типа чаще оказывают благоприятное действие, чем рецессивные. Это можно объяснить тем, что неблагоприятные доминантные мутации подвергаются более жесткому отбору - они уже в зиготе и на эмбриональных стадиях подвергаются элиминации, и отбором сохраняются лишь благоприятные. И так как само свойство доминирования гена эволюционирует под контролем отбора, то аллели дикого типа и оказываются более благоприятными для организма. Поэтому подбор в гибридной комбинации доминантных аллелей может скорее обеспечить гетерозис. Если бы гетерозис обусловливался простым набором доминантных аллелей, имеющихся в популяции, то этот набор было бы легко составить путем ряда скрещиваний и получить гетерозисные сочетания. Весьма возможно, что часть пород и сортов выводилась именно путем скрещивания и комбинаций линий, имеющих набор благоприятных доминантных аллелей. Но до сих пор не удавалось закрепить гетерозис в гибридной комбинации F 1 , т. е. получить не расщепляющиеся в F 2 формы.
Д. Джонсом еще в 1917 г. было предложено дополнение к изложенной гипотезе. Согласно Д. Джонсу, разные гены, дающие в сочетании гетерозис, находятся в пределах одной группы сцеплений: например, AbcdE - в одной, а соответственно aBCDe - другой гомологичной хромосоме. Именно это обстоятельство, с точки зрения Д. Джонса, затрудняет отбор в F 2 полностью гомозиготных форм по благоприятным доминантным генам, дающим сочетании гетерозис. Для осуществления такого сочетания доминантных аллелей необходимо, чтобы в этой паре хромосом AbcdE//aBCDe произошел минимум двойной кроссинговер, который привел бы к возникновению хромосомы с одними доминантными аллелями ABCDE. Малая вероятность одновременного осуществления такого события в нескольких хромосомах и является, по мнению Д. Джонса, причиной, затрудняющей закрепление гетерозиса. Гибрид же F 1 содержит все эти доминантные аллели, и поэтому у него наблюдается гетерозис.
Такова схема объяснения гетерозиса согласно второй гипотезе - гипотезе взаимодействия доминантных благоприятных генов, которую называют иногда также гипотезой набора кумулятивных доминантных аллелей, а сам гетерозис при этом - мутационным. Эта гипотеза в целом исходит из представления о простом суммировании эффекта доминантных аллелей с комплементарным действием.
Третья гипотеза исходит из того, что гетерозиготное состояние аллелей имеет превосходство над гомозиготным (АА < Aa > аа). Здесь можно допустить благоприятное трансположение аллелей дикого типа и мутантных аллелей, которое каким-то образом усиливает эффект действия генов. Указанное объяснение гетерозиса называют гипотезой сверхдоминирования.
Ни одна из трех гипотез не может считаться единственно правильной. Сейчас преждевременно отдавать предпочтение той или другой. Вероятно, они все окажутся правильными, но для разных случаев. Наверное, каждый из механизмов, предусматриваемых этими гипотезами, играет роль в определении гибридной мощности. Гетерозис является сложным явлением как по механизму возникновения, так и по проявлению его в онтогенезе.
Очевидно, окончательный вывод о генетических механизмах гетерозиса можно будет сделать лишь после того, как раскроется картина взаимодействия генов в генетической системе на биохимическом и молекулярном уровнях. Как мы уже знаем, подавление проявления мутаций с помощью супрессоров позволяет исправлять нарушения в действии гена и его аллелей. Возможно, что явление гетерозиса есть суммарный результат работы не основных генов, определяющих развитие признаков организма, а набора супрессирующих генов в генотипе. Важным моментом изучения гетерозиса становится исследование плазменных отношений.
Основной задачей использования гетерозиса в селекции является закрепление его, т. е. сохранение эффекта гетерозиса в процессе воспроизведения гибрида. Решение этой задачи мыслится в нескольких аспектах: во-первых, закрепление гетерозиса путем перевода гибридного организма с полового размножения на апомиктическое, что, по-видимому, возможно для некоторых растений; во-вторых, закрепление гетерозиса путем перевода диплоидного гибрида, проявляющего гетерозис, в полиплоидное состояние. В этом случае гетерозиготная комбинация генов будет сохраняться дольше.
У вегетативно размножающихся растений поддержание ценных гибридных комбинаций, полученных половым путем, осуществляется вегетативным размножением (черенками, прививками, клубнями и т. д.).
Существует ряд других путей поддержания гетерозиса в ряду поколений, однако все они еще недостаточно разработаны и проверена.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter .
Выберите правильный ответ:
1. Инбридинг – это:
1. скрещивание между родственными особями
2. скрещивание между неродственными особями
3. скрещивание между особями различного происхождения
2. Для каких растений инбридинг является нормальным способом размножения?
1. аутогамных
2. аллогамных
3. анеуплоидных
3. Генетическим следствием инбридинга является:
1. увеличение в потомстве доли гетерозиготных генотипов
2. увеличение в потомстве доли гомозиготных генотипов
3. появление в потомстве анеуплоидов
4.Гетерозис – это:
1. гомозиготизация гибридов F 1
2. ускорение роста, увеличение размеров, повышение жизнеспособности и продуктивности гибридов первого поколения по сравнению с родительскими формами
3. полиплоидизация гибридов F 1
5.У какого гибрида будет проявляться наибольший эффект «вредных» рецессивных генов?
1. АаВвссDdEeFf
6.Почему гетерозис затухает в последующих поколениях гибридов?
1. возрастает доля гомозиготных генотипов
2. возрастает доля гетерозиготных генотипов
3. возрастает доля анеуплоидных генотипов
7. Впервые гетерозисные межлинейные гибриды кукурузы получены:
1. Д. Джонсом
2. Г. Шеллом
3. В. Иогансеном
8. Почему у людей запрещены браки между близкими родственниками?
1. увеличивается вероятность проявления рецессивных вредных мутаций
2. увеличивается вероятность нарушений в мейозе
3. увеличивается вероятность хромосомных аббераций
9. Гетерозис – это
1. превосходство гибридов по жизнеспособности и продуктивности над обеими родительскими формами
2. полиплоидизация гибридов
3. гомозиготизация гибридов
10. Аутбридинг приводит к следующим последствиям:
1. повышается уровень гомозиготности потомства
2. повышается частота мутаций в потомстве
3. повышается уровень гетерозиготности потомства
11. Генетические механизмы закрепления гетерозиса:
1. различные формы бесполого размножения
2. семенное размножение
3. половое размножение
12. У каких растений инбридинг вызывает отрицательные последствия?
1. Аутогамных
2. Аллогамных
3. Анеуплоидных
13. Причиной снижения жизнеспособности организмов при инбридинге является:
1. проявление в потомстве летальных и других нежелательных генов
2. самонесовместимость
3. анеуплоидия
14. Популяции перекрестноопыляющихся культур состоят:
1. из гетерозиготных генотипов
2. из гомозиготных генотипов
15. Причина инцухт-депрессии заключается:
1. в реакции организма на изменчивость внешних условий
2. в проявлении отрицательных мутаций
3. в проявлении вредных рецессивных генов
16. Отрицательные последствия инбридинга:
2. возникновение инцухт-депрессии
3. повышение частоты мутаций
17. Максимальное проявление гетерозиса наблюдается:
18. Постепенное снижение жизнеспособности растений в инбредных поколениях, называется:
1. инверсией
2. интерференцией
3. инцухт-депрессией
19. Популяции самоопыляющихся культур состоят:
1. из гомозиготных генотипов
2. из гетерозиготных генотипов
20. Отрицательные последствия инбридинга:
1. повышение гетерозиготности потомства
2. повышение частоты мутаций
3. утрата ценных генов, имеющихся у исходной формы
21. Минимальное проявление гетерозиса наблюдается:
22. Гипотеза гетерозиготности (сверхдоминирования) объясняет гетерозис:
2. суммарным эффектом разнородных генетических процессов
3. взаимодействием доминантных генов
23. Инцухт-депрессия – это:
1. снижение частоты проявления доминантных аллелей
2. снижение гетерогенности популяции
3. постепенное снижение жизнеспособности растений от поколения к поколению
24. В результате самоопыления линии, гетерозиготной по одному аллелю (Аа), в потомстве можно выделить гомозиготные линии:
25. Благодаря чему неблагоприятные рецессивные гены в популяции
перекрестноопыляющихся культур находятся в скрытомсостоянии?
1. близкородственным скрещиваниям
2. свободным неродственным скрещиваниям
3. наследственной изменчивости
26. Чем обусловлена низкая выживаемость инбредных линий?
1. повышением частоты отрицательных мутаций
2. переходом летальных и полулетальных генов в гомозиготное состояние
3. повышением рекомбинационной изменчивости
27. Гипотеза доминирования объясняет гетерозис:
1. подавлением доминантными аллелями вредных рецессивных аллелей
2. гетерозиготностью аллелей одной и той же пары генов
3. благоприятным проявлением некоторых генов в гетерозиготном состоянии
28. Инбредные линии характеризуются:
1. гетерозиготностью
2. гомозиготностью
3. повышенной жизнеспособностью
29. В результате самоопыления линии, гетерозиготной по двум аллелям (АаВв), в потомстве можно выделить гомозиготные линии:
30. Что обусловливает гетерозиготность популяций перекрестноопыляющихся культур?
1. наследственная изменчивость
2. близкородственные скрещивания
3. неродственные скрещивания
31. У какого гибрида будет проявляться наибольший эффект «вредных» рецессивных генов?
2. АаВвссDd
32. При инбридинге мощность и урожайность линий:
1. повышается
2. снижается
3. сохраняется на одном уровне
33. Гипотеза доминирования объясняет гетерозис:
1. благоприятным проявлением некоторых генов в гетерозиготном состоянии
2. аддитивным действием благоприятных доминантных генов
3. гетерозиготностью аллелей одной и той же пары генов
34. Инбредное поколение принято обозначать буквой:
35. В результате самоопыления линии, гетерозиготной по трем аллелям (АаВвСс), в потомстве можно выделить гомозиготные линии:
36. У какого гибрида будет проявляться наименьший эффект «вредных» рецессивных генов?
1. АаВвссDdEe
37. Проявление гетерозиса в гибридных поколениях:
1. эффект гетерозиса повышается
2. эффект гетерозиса сохраняется
3. эффект гетерозиса ослабевает
38. Как называется скрещивание родственных особей?
1. инбридинг
2. аутбридинг
3. гетероплоидия
39. У какого гибрида будет проявляться наименьший эффект «вредных» рецессивных генов?
2. АаВвссDd
40. Гетерозис обусловлен:
1. мутационной изменчивостью
2. рекомбинационной изменчивостью
3. резким повышением гетерозиготности
41.Гетерозис закрепляется:
1. при семенном размножении
2. при вегетативном размножении
3. при половом размножении
42. Как называется скрещивание неродственных особей?
1. гетероплоидия
2. аутбридинг
3. инбридинг
43. У какого гибрида будет проявляться наименьший эффект «вредных» рецессивных генов?
1. АаВвссDdEeFf
44. Число гомозиготных генотипов для n локусов соответствует:
45. Явление гетерозиса было открыто:
1. С.Г. Навашиным
2. И.Г. Кельрейтером
3. С.С. Четвериковым
46. Принудительное самоопыление у перекрестноопыляющихся организмов приводит:
1. к вспышке гетерозиса
2. к снижению жизнеспособности
3. к повышению урожайности
47. У какого гибрида будет проявляться наибольший эффект «вредных» рецессивных генов?
1. АаВвссDdEe
Установите соответствие:
48. Соотношение генотипов при самоопылении гетерозиготы Аа в 1-м
1.1. Под термином гетерозис в широком смысле слова понимают все положительные эффекты, ведущие к превосходству гибридов первого поколения (F 1) над родительскими формами.
Гетерозис (от греч. heteroiosis - изменение, превращение), «гибридная сила», ускорение роста к увеличение размеров, повышение жизнестойкости и плодовитости гибридов первого поколения по сравнению с родительскими формами при различных скрещиваниях как животных, так и растений. Гетерозис в полной мере проявляется в первом гибридном поколении. Во втором и последующих поколениях Г. обычно затухает. У вегетативно размножающихся растений, например у картофеля, сахарного тростника и др., гетерозис передается вегетативному потомству стойко, так как все растения клона генотипически соответствуют исходной материнской особи.
Преимущества гетерозисных гибридов : высокое качество семян, повышенный урожай, устойчивость к многим болезням, очень ранние сроки созревания.
Недостатки гетерозисных гибридов : высокая цена семян, высокие требования к агротехнике, высокая генетическая однородность гибридов приводит к сильному распространению заболеваний, невозможность воспроизведения и собора семена.
Селекция гетерозисных гибридов имеет большое значение для сельскохозяйственного производства. Эти гибриды часто по урожайности превышают обычные свободноопыляющиеся сорта на 30% и выше. В некоторых случаях гетерозисный эффект достигает 50%. Широко используется явление гетерозиса в селекции кукурузы, сорго, под солнечника, томата, тыквы, огурца, арбуза, лука, капусты, сахарной свеклы, декоративных растений. Его начат применять также в селекции риса, пшеницы, хлопчатника.
1.2. ФАКТОРЫ, ОБУСЛОВЛИВАЮЩИЕ ГЕТЕРОЗИС
Долгое время явление гетерозиса пытались объяснить отдельными генетическими факторами. Возникло несколько гипотез.
Так, гипотеза сверхдоминирования объясняет проявление гетерозиса гетерозиготным состоянием гибридов.
Вторая классическая гипотеза о проявлении гетерозиса, так называемая гипотеза доминирования , исходит из того, что не гетерозиготное состояние само по себе, а вызванное скрещиванием накопление доминантных аллелей продуктивности приводит к гетерозису.
Многие исследования, проведенные после появления указанных гипотез, показали, что причины проявления гетерозиса комплексные . Очевидно, гетерозис обусловлен физиологической сбалансированностью процессов обмена веществ , что достигается гетерозисными гибридами легче, чем гомозиготными формами.
На основании этого была выдвинута концепция генетического баланса . Оказалось, что и ядерно-плазматические взаимодействия могут влиять на обусловленную генами хромосом сбалансированность гибридов, особенно если в гибридном идиотипе присутствуют генетически различные митохондрии, что может привести к усилению дыхания и энзиматической активности. Таким образом, гетерозис определяется не только генотипической, но и плазматической конституцией гибридов.
1.3. ТИПЫ ГИБРИДОВ, ИСПОЛЬЗУЕМЫХ В ПРОИЗВОДСТВЕ
Различают следующие типы гибридов производственного использования:
1) межлинейные:
Простые - от скрещивания двух самоопыленных линий;
Трехлинейные - от опыления простого межлинейного гибрида пыльцой самоопыленной линии;
Двойные - от скрещивания двух простых межлинейных гибридов;
Сложные межлинейные гибриды - получаемые с участием более четырех самоопыленных линий;
2) сортолинейные:
Простые - от опыления сортов пыльцой линии;
Сложные - от опыления сорта пыльцой простого межлинейного гибрида;
3) линейносортовые - от опыления простого гибрида пыльцой сорта;
4) межсортовые гибриды - от скрещивания двух сортов;
5) гибридные (синтетические) популяции - получаемые путем смешения семян простых гибридов и других компонентов» их свободного переопыления.
Наиболее высокую прибавку урожая дают гибриды, полученные с участием самоопыленных линий.
1.4. ОБЩАЯ СХЕМА СЕЛЕКЦИИ ГЕГЕРОЗИСНЫХ ГИБРИДОВ
Гетерозисная селекция состоит из нескольких этапов. Селекционная работа начинается с выбора исходного материала, из которого создают самоопыленные линии. Затем изучают комбинационную способность этих линий, а наиболее пригодные к комбинированию используют в качестве родительских форм для создания простых, двойных и других типов гибридов и для составления гибридных популяций.
-
 17 апреля 2015Tom Clancy’s The Division - Дополнение «Под землей»
17 апреля 2015Tom Clancy’s The Division - Дополнение «Под землей» -
 17 апреля 2015Заполнение журналов-ордеров
17 апреля 2015Заполнение журналов-ордеров







